












|
HUD |
| Submitted By PrinzAdium |
DescriptionLooks great with:»Service Extensions« service icons (http://adiumxtras.com/index.php?a=xtras&xtra_id=3145) »Candyball« status icons (http://adiumxtras.com/index.php?a=xtras&xtra_id=2010) »Twitterrific« status icons (http://adiumxtras.com/index.php?a=xtras&xtra_id=4103) Note: 73% opacity but maybe up to 90% looks also good. Changesimproved colorsbug fix Images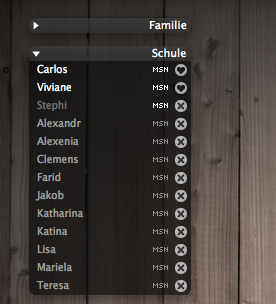 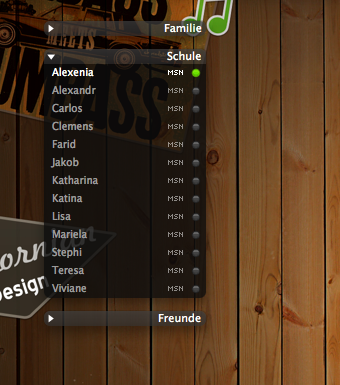 CommentsYou can reply to individual comments by clicking the "Reply" link next to each. # by zazzakakashi on 12/15/07 at 16:35:45I love this contact window style! I cant wait for someone to come up with the message style and my life would be pretty complete LMAO ! Keep up the great work!
# by m1kee on 12/31/07 at 09:42:32Oh wow I LOVE this :D 5 STARS! And where'd you get the desktop background? I waaant it :)
# by PrinzAdium on 01/14/08 at 13:52:58# by rikibrucelee on 01/19/08 at 13:23:00Wonderfull!!
I like the wallpaper of your second screenshot (wood & posters). how can I find it in hires version (1920x1200 or 1650x1080)? Thanks # by PrinzAdium on 01/19/08 at 15:29:04Thank you! The wallpaper has the dimensions 2500x1600. I think you can use it as it is.
Best # by PrinzAdium on 02/01/08 at 15:19:21# by Lockyssimo on 02/17/08 at 13:38:22Someone could give me the link status icons heart(love), please!
# by ehmjay on 04/26/08 at 01:01:05I love this theme so much, but I was wondering if anyone has some suggestions for chat styles to match up with it.
# by PrinzAdium on 04/26/08 at 08:09:39Thank you for the comments!
Chat Style? I don't know I'm just trying to add a blur effect to the window. # by m0nk3y on 10/16/08 at 14:58:57can any one recommend a good message style to go with this? I have mine setup similar to the second pic, but i can't find a good mesage style that ties in with it. Thanks!
# by jayrobinson on 10/31/08 at 00:03:43The best message style to use with this is Ethereal, which allows for a transparent background and the smoke appearance. http://www.adiumxtras.com/index.php?a...tra_id=2141
# by haiggy on 09/10/09 at 07:59:01What font is that?
My Adium list doesn't look like this at all when I change my theme and install it correctly. Every font for every theme is always Lucida Grande... what the heck? # by pixelologist on 10/06/09 at 05:49:20I love this theme with the 1337 message style (saturated style). Only thing - and I don't know if this is an issue peculiar to this theme or if it happens with Adium generally - is that when I use the Group Bubbles variation, every once in a while my contact list suddenly acquires an unnecessary vertical scroll bar on the right. The only way to get rid of it is to open prefs, change to Regular Window and then back to Group Bubbles. Is there any way to prevent that from happening? Or to deselect a contact name while in Group Bubbles mode?
# by thesr5 on 10/09/09 at 06:01:08This may be a stupid question and may not be able to be done, but i see in your pictures that there is no toolbar (at all)... how do i remove mine??
# by unchoian on 07/24/10 at 10:03:54very neat and beautiful : )
i am using it right now :D thanks so much! wanna ask u abt the wooden wallpaper :P # by LcTheSecond on 10/21/10 at 12:30:59Hey man, is it possible (with the current theme) to leave the background transparent ???
If so, how to do it? If not, could you add the option to do it ? is it even possible ? # by LcTheSecond on 10/21/10 at 12:33:14Ps. How do you leave "Msn" instead of the icons ?? Like on the print screen.
Post a New CommentYou must be logged in to post comments. |

# by Dan1jel on 11/21/07 at 17:50:45
btw can u tell me how much Opacity u have?